Velika enciklopedija nafte i gasa. Molekularna struktura materije
Nau?nici su dugo vremena stvarali teorije i modele koji bi pomogli da se objasne osnovna svojstva supstanci i materije koje ?ine svijet oko nas.
U toku istorije izvedeni su mnogi eksperimenti, eksperimenti; otkriveni su novi zakoni i fizi?ka i hemijska svojstva materijala. To je uglavnom bilo zbog otkri?a pojmova "molekula" i "atomsko-molekularna struktura materije". Razgovarajmo o njima detaljnije u ovom ?lanku.
Pojava koncepta "atomsko-molekularne strukture materije"
Jo? u doba anti?ke Gr?ke pojavila se ideja da se sve na svijetu sastoji od najsitnijih dijelova. Grci su te ?estice nazivali molekulama i atomima. Autor ove hipoteze bio je Demokrit, koji je kasnije postao osniva? atomisti?ke teorije. Ali ovo znanje u to vreme nije se mnogo razvilo sve do 17. veka. Sva istra?ivanja materijala opet su dovela do ?injenice da su mnoge tvari sastavljene od molekula, ?ija su strukturna jedinica atomi.
Kasnije su nau?nici po?eli da dolaze do zaklju?ka da se jedna vrsta minerala sastoji, na primer, od 38% gvo??a i 62% kiseonika, a svaki od uzoraka ?e pokazati takav hemijski sastav. Ali ako uzmemo drugo tijelo s odli?nim svojstvima, onda ?e analiza atomske i molekularne strukture tvari pokazati da se sastoji od 60% ?eljeza i 40% kisika.
Dakle, unato? ?injenici da jo? uvijek nije postojao odre?eni koncept, u toku istra?ivanja po?ela se pojavljivati teorija da je svaka supstanca druga?iji skup molekula i atoma koji odre?uju njena osnovna svojstva.
Razvoj koncepta "molekularne strukture materije"

Pojam "molekula" prvi je uveo 1811. godine talijanski fizi?ki hemi?ar Amadeo Avogadro. Upravo je on postao osniva? teorije atomske i molekularne strukture.
Potvrda ove teorije pojavila se tek 1860-ih, kada je ruski hemi?ar A. M. Butlerov formirao i uspio objasniti molekularnu teoriju strukture materije. U skladu sa njegovim radovima, svojstva bilo koje supstance odre?uju se time kako su atomi u molekulima me?usobno povezani, njihovom interakcijom. Nau?nik je iznio hipotezu da je molekul mikro?estica materijala, koja se sastoji od atoma, koja mo?e postojati nezavisno.
Koncept molekula uspostavljen je zahvaljuju?i radu jo? jednog ruskog nau?nika. Govorimo o M. V. Lomonosovu.
U razvoju atomisti?ke doktrine u?estvovali su mnogi nau?nici iz celog sveta: J. Maxwell, L. Boltzmann, J. Gibbs, R. Clausius, J. Dalton, D. I. Mendeleev, V. Roentgen, A. Becquerel, J. Thomson , M .Planck i mnogi drugi. Doprinos ovih ljudi molekularnoj fizici i hemiji je neprocjenjiv.
Su?tina molekularne teorije strukture materije

Na osnovu niza vje?bi tokom godina, izvedena je molekularna teorija strukture materije. Postoji nekoliko osnovnih odredbi ove teorije, odnosno tri osnovne tvrdnje koje su vi?e puta dokazane kroz laboratorijska istra?ivanja:
- Svako tijelo se sastoji od najmanjih ?estica - molekula i atoma, koji se tako?er sastoje od manjih elemenata. Struktura svih supstanci je diskontinuirana.
- Atomi i molekuli su u stalnom kretanju haoti?ne prirode.
- Sve tvari me?usobno djeluju na osnovu elektromagnetnih sila privla?enja i odbijanja.
Utemeljenje teza molekularne kineti?ke teorije
Prva potvrda odredbi teorije je Brownovo kretanje, koje je 1827. godine otkrio poznati botani?ar R. Brown. Razlog za ovaj fenomen je haoti?no kretanje molekula u razli?itim smjerovima, koje nastaje kao rezultat njihovih sudara.
Druga potvrda ove teorije bit ?e bezbrojni eksperimenti s procesom difuzije, odnosno sposobno??u jedne supstance da prodre u drugu. Vrhunski primjer takvog iskustva iz svakodnevnog ?ivota je parfem ili bilo koja aromati?na supstanca. Ako se takva supstanca stavi u prostoriju, nakon odre?enog vremena aroma ?e se raspr?iti po cijeloj njenoj povr?ini.
Atom i molekul
Danas su ovi pojmovi ve? potkrijepljeni i izvedeni ta?no i razumno. Jednostavno re?eno, atom je kemijski nedjeljiva ?estica bilo koje tvari ili materijala koji ?ini molekul. A molekul je i najmanja ?estica ne?ega, ali bitno je da ona postavlja osnovna svojstva tijela. Molekularna struktura tvari je okvir odre?enog materijala, u ?ijim spojevima se nalaze molekuli.

Ako materijal karakterizira molekularna kristalna re?etka, tada on, u pravilu, ima nisku tvrdo?u, lako se rastopi; takva supstanca ?e biti hlapljiva ili rastvorljiva u vodi, elektri?na energija ne provodi.
Nauka koja prou?ava silu interakcije izme?u ovih ?estica i molekularne strukture tijela naziva se molekularna fizika. Ovdje se istra?uju razli?ita svojstva tijela u razli?itim agregacijskim stanjima.
Koje supstance imaju molekularnu strukturu?
Molekularna veza je op?enito slaba i prevladava u organskim tvarima. Mnoge nama poznate supstance imaju molekularnu strukturu. Na primjer, voda (H 2 O), vodonik (H 2), hlor (Cl 2), uglji?ni dioksid (CO 2), kisik (O 2), etanol ili etil alkohol (C 2 H 5 OH), organski polimeri i mnogo drugih.

Drugim rije?ima, tvari koje imaju molekularnu strukturu su uglavnom plinovi. U njima su molekuli udaljeni jedan od drugog i slabo me?usobno djeluju. Bliska veza izme?u ?estica tvari stvara ?vrste tvari. Jedina te?nost koja ima molekularnu strukturu je Br 2 . Ova supstanca je vrlo isparljiva.
Nemetali s molekularnom strukturom uklju?uju ?vrste tvari kao ?to su I 2, P 4, S 8. Ovi materijali su topljivi i mogu sublimirati.
Molekularna struktura, tj. hemijski sastav i na?in povezivanja atoma u molekulu ne odre?uju jednozna?no pona?anje polimernog materijala izgra?enog od makromolekula. Svojstva polimera, posebno u stanju kristalne faze, zavise od njihove supramolekularne strukture, tj. metoda pakovanja makromolekula u prostorno izdvojene elemente, veli?ina i oblik takvih elemenata i njihov relativni polo?aj u prostoru. Drugim rije?ima, pod supramolekularnom strukturom se podrazumijevaju slo?eni agregati velikog broja makromolekula, koji nastaju kao rezultat djelovanja me?umolekularnih sila.
?vrsta i te?na agregiraju?a stanja su tipi?na za polimere, karakterizirana oscilatornim i rotacijskim kretanjem ?estica i malim razmacima izme?u ?estica. Polimeri ne postoje u gasovitom stanju, jer Da bi se makromolekule razdvojile na velike udaljenosti, potrebno je savladati jake me?umolekularne interakcije lan?anih makromolekula, za koje su potrebne energije uporedive sa energijama hemijskih veza u polimernom lancu, tj. dolazi do razgradnje polimera.
Fazno stanje je odre?eno redoslijedom u rasporedu molekula. Postoje dvije vrste faznog stanja: amorfno i kristalno. Stanje amorfne faze karakteri?e poredak kratkog dometa na udaljenostima od 10-15?. Termodinami?ki stabilno stanje je izotropno, iako su u amorfnim polimerima mogu?e lokalne ure?ene formacije fluktuacijskog karaktera. Jedan od prvih modela strukture amorfnog stanja polimera je Karginov model "prskanja". Pretpostavljeno je da za optimalno pakovanje dugolan?anih molekula u amorfnim polimerima postoje ure?ena podru?ja u obliku intermolekularnih paketa (IMP) formirana od paralelno orijentiranih susjednih makromolekula pro?irene pro?irene konformacije. Shodno tome, glavni strukturni element linearnih polimera sa fleksibilnim lancem u amorfnom stanju nije sama makromolekula, ve? MMP ili neka druga supramolekularna struktura, u kojoj pojedina?ni makromolekuli gube svoju individualnost.
Stanje kristalne faze karakteri?e trodimenzionalni poredak dugog dometa na udaljenostima od 1000?. Ovo stanje se odlikuje anizotropijom svojstava, skokovima svojstava na granici faze. U kristalnim polimerima gotovo uvijek postoji dio amorfne faze; ?esto se susre?u defekti i dislokacije. Pote?ko?e u dobivanju polimernih kristala i karakteristike kristalnog stanja makromolekula povezane su s raznim supramolekularnim strukturama koje jo? uvijek postoje u amorfnom stanju.
Pored amorfnog i kristalnog, poznato je i mezofazno srednje te?no-kristalno stanje polimera. Odlikuje se stalnom stabilnom anizotropijom nekih fizi?kih svojstava. U obi?nim izotropnim te?nostima mo?e nastati privremena „indukovana“ anizotropija pod uticajem elektri?nog polja, mehani?kih uticaja itd. Nasuprot tome, u polimerima, zbog male pokretljivosti makromolekula i dugog vremena relaksacije, „indukovana“ anizotropija opstaje. na neodre?eno, tj. je odr?iv.
Supramolekularna struktura amorfnih polimera
Najpotpunija ideja o procesima formiranja supramolekularnih struktura i njihovim tipi?nim oblicima mo?e se dobiti ako se prati cijeli proces formiranja strukture. Postoje dva na?ina formiranja struktura. Ako su makromolekule dovoljno fleksibilne, mogu smotati sferne ?estice (zavojnice), takozvane globule.
Me?usobni raspored pojedinih dijelova lan?ane makromolekule unutar takve globule je po pravilu nasumi?an, a gotovo svaki polimer pretvoren u oblik globula je u amorfnom stanju. Na primjer, polietilen, poliamidi.
U vrlo razrije?enim otopinama, velika ve?ina polimera je u obliku takvih zavojnica. Naj?e??i na?in za dobivanje polimera u globularnom stanju je isparavanje otapala iz otopina na najni?im mogu?im temperaturama. Makromolekule brojnih proteina su tako?er u globularnom stanju. Kuglasta struktura je korisna samo za transport polimerne supstance u otopljenom stanju, ?to je vrlo va?no za biolo?ke procese. Za sve ostale slu?ajeve, to zna?i gubitak glavnih svojstava povezanih sa linearnom lan?anom strukturom makromolekula.
Sposobnost globularnih polimera da formiraju slo?enije strukture vrlo je ograni?ena. Ako je polimer monodisperzan, tj. sve njegove makromolekule su iste, a zatim se globule sla?u u strukturu sa zbijenim lopticama. Tako nastaju monokristali globularnih proteina. Na primjer, virus mozaika duhana.
Globule nastaju kao rezultat vi?ka sile intramolekularne interakcije nad silama me?umolekularne interakcije.
Osim toga, da bi se promijenio iz izdu?enog u globularni oblik, molekularni lanac mora imati dovoljno veliku fleksibilnost da bi se mogao savijati.
Rice. 1. Molekul superljepila
Ako je polimer u visoko elasti?nom stanju, tada se pojedina?ne ?estice globule mogu spojiti u jednu ve?u ?esticu. Pojavljuju se globule koje sadr?e vi?e (desetak, stotina, hiljada) ?estica. Ovaj proces se zavr?ava ili raslojavanjem sistema, ili stabilizacijom formiranih velikih globula, zbog oblaganja njihove povr?ine ne?isto?ama ili njihovog preure?ivanja u linearne strukture. Sli?ni procesi se de?avaju tokom polimerizacije. U zavisnosti od uslova polimerizacije, odre?ene konformacije mogu biti termodinami?ki povoljnije. Stoga je od istog polimera mogu?e dobiti mnogo polimera razli?ite fizi?ke strukture, ?iji ?e ekstremni tipovi biti globularni i fibrilarni.
Poznato je da postoje odvojeni produ?eni linearni lanci polimernih supstanci. Pro?ireni makromolekularni lanci formiraju linearne agregate - lan?ane pakete. Tipi?na karakteristika ovih formacija je da njihove du?ine znatno prema?uju du?inu pojedina?nih lanaca. Svaki od ovih paketa uklju?uje desetine, stotine pojedina?nih makromolekula. Ovi snopovi lanaca su nezavisni strukturni elementi, od kojih se zatim grade slo?enije strukture.
Model praska je nedavno pro?ao kriti?ku reviziju. Weinstein, koji je prou?avao difrakciju X-zraka na amorfnom polimeru, do?ao je do zaklju?ka da struktura takvog polimera ne mo?e biti snop. Du?ina sekcija paralelizma polimernih molekula, po njegovom mi?ljenju, nije velika i pribli?no je jednaka njihovoj ?irini. Ovi dijelovi postepeno prelaze jedan u drugi, unutar njih i u prijelaznim zonama izme?u njih nastaju "kosi" kontakti molekula. Osim toga, model praska je u suprotnosti s glavnim odredbama kineti?ke teorije visoke elasti?nosti, ?to je dobro potvr?eno eksperimentom.
Yeh je predlo?io drugi model za supramolekularnu organizaciju amorfnog polimera. On je sugerisao da amorfno stanje polimera karakteri?e prisustvo ure?enih regiona - domene("zrna") formirana od makromolekula sa naboranim konformacijama.
Domeni su me?usobno povezani pomo?u lanaca petlji. Interdomenske regije se sastoje od karika nasumi?no raspore?enih lanaca, a tako?er uklju?uju prolazne lance i slobodne krajeve lanaca koji nisu uklju?eni u domene.
Postoje tri mogu?a tipa domena: presavijeni (rebrasti), snop?asti (micelarni) i globularni. Posljednja dva tipa se dobro sla?u sa teorijom praska i globularom. Ovo pokazuje zasluge teorije domena kao op?enitije i objedinjuju?e.
Postojanje domena u topljenju polimera varira. Domeni - skup me?umolekularnih veza, nastaju i kolabiraju pod uticajem toplotnog kretanja. ?to je temperatura vi?a, ?ivotni vijek domene je kra?i i njegova veli?ina je manja. Sa smanjenjem fleksibilnosti lanaca i njihove pravilnosti, smanjuje se i tendencija formiranja domena.
Nije uvijek mogu?e otkriti domene u topljenjima polimera sa krutim nepravilnim lancima. Smatra se da se u takvim uslovima ne formiraju, a lanci imaju oblik statisti?kih zavojnica-globula. Zavojnice prodiru jedna u drugu, formiraju?i direktne kontakte.
Za razliku od Yehe, Arisakov, Bakeev i Kabanov, koriste?i Yehine modele, smatraju da se amorfni polimer sastoji od gusto zbijenih fibrila. Svaka fibrila se sastoji od presavijenih domena povezanih prolaznim lancima. Ali eksperimentalni podaci ne dopu?taju nam da smatramo fibril kao glavni oblik supramolekularne organizacije amorfnog polimera.
Tako?e je predlo?en model klastera koji obja?njava supramolekularnu organizaciju amorfnih polimera.
Klasteri su podru?ja u kojima postoji gu??e pakiranje molekula ili ?estica, kao i njihov ure?eniji raspored u odnosu na glavnu, labaviju i neure?eniju masu tvari. Stoga gustina klastera ne?to prema?uje prosje?nu gusto?u polimera u cjelini. Ali u pore?enju sa kristalima, klasteri su manje ure?eni i manje gusto zbijeni regioni. U tom smislu, postoje dvije vrste klastera:
1. kristalno - klasteri u kojima se pod odre?enim uslovima mo?e posti?i ure?eniji raspored makromolekula. Oni su u stanju da kristalizuju.
2. anti-kristalni- klasteri koji se u osnovi ne kristali?u.
Amorfni polimer je skup antikristalnih klastera okru?enih manje ure?enim i labavim podru?jima. Posljedi?no, gustina amorfnih polimera je direktno proporcionalna zapreminskom udjelu klastera. To se poklapa s takvom eksperimentalnom ?injenicom kao ?to je pove?anje gusto?e amorfnih polimera tokom njihovog ?arenja. ?arenje dovodi do pove?anja broja antikristalnih klastera, pove?anja prosje?ne veli?ine ovih formacija i ure?enijeg rasporeda polimernih lanaca u njima.
Amorfni polimeri imaju nasumi?nu orijentaciju svojih polimernih lanaca, dok kristalni polimeri formiraju visoko ure?enu kristalnu strukturu u amorfnoj matrici (slika 2). Termin polukristalni polimeri koristi se za polimere koji sadr?e i kristalne i amorfne regije.

Rice. 2. Amorfni polimeri
Posebnost modela klastera je da mu nedostaje regulisana priroda rasporeda lanaca unutar klastera (odre?ena je hemijskom strukturom polimera, njegovom molekulskom te?inom). Klaster se mo?e sastojati od kako makromolekula sa savijenom konformacijom, tako i nesavijenih elemenata polimernih lanaca koji ne formiraju nabore. Druga karakteristika je fluktuacijski karakter klastera.
Gore opisane organizacije makromolekula u amorfnim polimerima samo su najjednostavniji prikazi oblika ure?enja makromolekula. Oni su va?ni jer su prva faza u procesima organizacije makromolekula, bez kojih su nemogu?e razli?ite supramolekularne strukture kristalnih polimera.
Supramolekularna struktura kristalnih polimera
Fazno stanje polimera ili sposobnost polimera da kristalizuju zavisi od mnogih uslova: temperature i brzine kristalizacije, termi?ke istorije, prisustva stranih supstanci. U zavisnosti od uslova kristalizacije, mogu se dobiti mnogi morfolo?ki oblici kristalnih struktura ?ak i za isti polimer. Raznolikost supramolekularnih struktura u kristalnim polimerima je posljedica fleksibilnosti i strukture dugog lanca polimera.
Jedna od karakteristika kristalnog stanja polimera je prisustvo u njima zna?ajnog udjela poreme?aja - "udio amorfne faze". Stoga se posebna pa?nja poklanja procesima kristalizacije.
Kada se rastvara? ukloni iz razrije?ene otopine polimera, u slu?aju dovoljno jake intermolekularne interakcije, makromolekule se mogu povezati u paket. Pakovanje je primarna supramolekularna struktura.
Ako je pakiranje formirano od pravilnih fleksibilnih makromolekula, tada ?e u njemu, pod odgovaraju?im termodinami?kim uvjetima, do?i do kristalizacije, odnosno, makromolekule ?e se rasporediti tako da formiraju prostornu re?etku. Kristalizirano pakovanje ima me?uprostor i povr?insku napetost karakteristi?nu za kristalnu supstancu. Me?utim, pojava vi?ka povr?inske energije trebala bi biti zna?ajna u tako tankim i dugim formacijama.
Kao rezultat, kristalno pakovanje stje?e sposobnost savijanja u trake sa manjom povr?inskom gustinom. Traka je energetski povoljnija forma. Preklapanje paketa u trake doga?a se spontano u smjeru smanjenja slobodne energije F<О путем многократного поворота пачки на 180°. Лента - вторичная надмолекулярная структура при пластинчатом механизме кристаллизации. Существование складчатых структур было впервые обнаружено и детально исследовано Келлером на примере полиэтилена и полиамидов.
Formiranje sekundarnih struktura u kristalnom polimeru ne prestaje na formiranju vrpci. Zahtjev za smanjenjem povr?inske napetosti dovodi do savijanja "traka" u ravne formacije, tj. u letvicama. Takve plo?e se formiraju spajanjem pojedina?nih "traka" svojim ravnim stranama, ?to dovodi do daljeg smanjenja povr?ine. Za linearne polimere iz slo?enih lanaca tipi?ni su lamelarni kristali, koji su prili?no savr?eni pri niskim brzinama kristalizacije (monokristali polietilena).

Rice. 3. Pruge pokazuju rast polietilenskih kristala
Osim lamelarnog mehanizma formiranja monokristala, postoji jo? jedna vrsta struktura, koju karakterizira prisustvo fibrilarnih elemenata.
Najva?niji prakti?ni primjer dobijanja kristala u kojima lanci u velikoj mjeri zadr?avaju ispravljene konformacije je kristalizacija pri hla?enju taline uz istovremenu primjenu visokih naprezanja. Nastale strukturne forme, nazvane "?i?-kebab", karakterizira prisustvo dugog fibrilarnog sredi?njeg trupa. Na ovom ?tapu rastu lamele u popre?nom smjeru, u kojem su lanci u presavijenim konformacijama.
Uz lamelarni mehanizam formiranja monokristala, postoji jo? jedan tip pojavljivanja niza vi?ih supramolekularnih struktura. Kod lamelarnog tipa najjednostavniji strukturni element je plo?a traka, dok kod fibrilarnog tipa takvo savijanje nema i snopovi se nalaze du? fibrile. Jedan te isti polimer mo?e kristalizirati i lamelarnu i fibrilarnu.
Slika formiranja monokristala je ograni?avaju?a slika procesa kristalizacije. To implicira neograni?ene mogu?nosti za nastanak neravnote?nih stanja kristalnih polimera. Kada se kristalizacija kasni u fazama formiranja vrpci, plo?a i fibrila, to dovodi do stvaranja sferulitnih struktura - naj?e??eg elementa velikih struktura.
To su agregati kristala sa jednim centrom i radijalnom orijentacijom kristala u odnosu na centar.
To su tipi?ne polukristalne formacije dobivene u realnim uvjetima za formiranje odljevaka, filmova i drugih polimernih proizvoda na bazi kristaliziraju?ih makromolekularnih spojeva gotovo svih klasa.

Proizvodi od polistirena. Fotografija: Pat Hayes
Sferuliti nisu termodinami?ki povoljni, ali su kineti?ki po?eljniji. Veli?ine sferulita mogu varirati u ?irokom rasponu - od desetina mikrona do nekoliko milimetara ili vi?e. Mali sferuliti pokazuju sposobnost agregacije sa formiranjem veoma dugih ?estica nalik vrpci.
Trake napravljene od sferulita karakterizira anizotropija opti?kih svojstava zbog radijalne asimetrije njihove strukture. Promjena orijentacije se doga?a u svakom radijalnom smjeru, ?to se manifestira pojavom uzorka naizmjeni?nih svijetlih i tamnih prstenova.

Grafitni sferuliti vi?eni pod elektronskim mikroskopom
Osim radijalnih, postoje i prstenasti sferuliti, koji se odlikuju ?injenicom da se na ?aru malte?kog kri?a nadogra?uje izra?en sistem naizmjeni?nih tamnih i svijetlih prstenova.
U procesu kristalizacije na sukcesivno ni?oj temperaturi mogu se dobiti razli?ite me?ufaze od poliedarskih lamela do visoko razgranatih dendrita.
Dendriti su razgranati kristali, koji ponekad izgledaju kao drvo (od gr?kog za "drvo"). Sve grane dendrita su kristalografski povezane jedna s drugom.

Supramolekularne strukture u polimerima prou?avaju se elektronskom mikroskopijom, neutronskom difrakcijom, difrakcijom rendgenskih zraka, raspr?ivanjem svjetlosti, dvolomom itd. Posebno su metodom difrakcije neutrona malog snopa odre?eni radijusi rotacije makromolekula, koji se poklapaju sa neporeme?ene veli?ine Gaussovih zavojnica i nepro?irenih "pakovanja". Trenutno su poznati alternativni strukturni modeli: preklapaju?i statisti?ki kalemovi (OSC); statisti?ki slo?ene makromolekule (domene) itd. Konkretno, model PSC koji je predlo?io Flory omogu?io je da se teorijski potkrijepi koncept isprepletenosti i postao je osnova za statisti?ke teorije strujanja i termodinami?kih svojstava koncentriranih polimernih otopina. Rezultati kompjuterske simulacije konformacija metodom Monte Carlo tako?er su potvrdili PSC, koji se odlikuje prili?no zbijenim pakiranjem.
Metode za prou?avanje strukture polimera mogu se podijeliti u dvije grupe. Prvi uklju?uje vizuelne metode: opti?ke i elektronska mikroskopija, u kojima je kori?tena valna du?ina (izvora svjetlosti ili snopa elektrona) mnogo manja od dimenzija strukturnih elemenata (makromolekula ili njihovih agregata).
Druga grupa uklju?uje interferencija-difrakcija metode: Difrakcija rendgenskih zraka, difrakcija elektrona i neutrona, raspr?ivanje svjetlosti. Ove metode koriste elektromagnetne oscilacije sa talasnom du?inom koja je uporediva sa veli?inom prou?avanih strukturnih elemenata. Na primjer, jedna od naj?e??ih metoda - analiza difrakcije rendgenskih zraka - zasniva se na fenomenu difrakcije rendgenskih zraka sa l=0,5-2,5?. Ako rendgenski snop padne na kristale ?ije su linearne dimenzije uporedive, tada je mogu?e procijeniti period identiteta, utvrditi relativni polo?aj razli?itih ravni kristalne re?etke, procijeniti stepen kristalnosti, veli?ine kristala i njihovu orijentaciju. .
Pomo?u elektronske mikroskopije mogu se posmatrati pojedina?ne makromolekule i njihovi agregati. Ovom metodom dobijeni su glavni tipovi supramolekularnih struktura prikazanih na slikama: fibrilarni kristali, monokristali i sferuliti. Fini detalji strukture sferulita mogu se prou?avati samo pomo?u elektronskog mikroskopa.
Prisustvo sferulita uti?e na mehani?ka (?vrsto?u) i druga svojstva polimera. Na primjer, neprozirnost polietilena, najlona i drugih kristalnih polimera je posljedica prisustva sferulita. Raznolikost supramolekularnih struktura glavni je razlog posebnih svojstava kristalnih polimera.
Jedan od glavnih razloga interesovanja za fiziku makromolekula je njeno kori??enje za razumevanje tajni ?ive prirode, za razumevanje molekularnih osnova pona?anja biolo?kih sistema. Napredak u razumijevanju mehanizma ?ivotnih procesa nemogu? je bez primjene fizi?kih i hemijskih ideja i metoda za prou?avanje biolo?kih procesa na molekularnom nivou.
(molekularna struktura), me?usobni raspored atoma u molekulima. U toku hemijskih reakcija, atomi u molekulima reaktanata se preure?uju i nastaju nova jedinjenja. Stoga je jedan od temeljnih kemijskih problema razjasniti raspored atoma u po?etnim spojevima i prirodu promjena u formiranju drugih spojeva iz njih.Prve ideje o strukturi molekula bile su zasnovane na analizi hemijskog pona?anja materije. Ove ideje su postale slo?enije kako se akumuliralo znanje o hemijskim svojstvima supstanci. Primena osnovnih zakona hemije omogu?ila je odre?ivanje broja i vrste atoma koji ?ine molekul datog jedinjenja; ova informacija je sadr?ana u hemijskoj formuli. Vremenom su kemi?ari shvatili da jedna hemijska formula nije dovoljna da se precizno okarakteri?e molekul, jer postoje molekuli izomera koji imaju iste hemijske formule, ali razli?ita svojstva. Ova ?injenica navela je nau?nike na ideju da atomi u molekulu moraju imati odre?enu topologiju, stabilizovanu vezama izme?u njih. Ovu ideju prvi je iznio 1858. njema?ki hemi?ar F. Kekule. Prema njegovim idejama, molekul se mo?e prikazati pomo?u strukturne formule, koja ukazuje ne samo na same atome, ve? i na veze izme?u njih. Interatomske veze tako?e moraju odgovarati prostornom rasporedu atoma. Faze u razvoju ideja o strukturi molekula metana prikazane su na Sl. 1. Struktura odgovara modernim podacima
G : molekula ima oblik pravilnog tetraedra, u ?ijem se sredi?tu nalazi atom ugljika, a na vrhovima su atomi vodika.(6.58 Kb)Takve studije, me?utim, nisu govorile ni?ta o veli?ini molekula. Ove informacije su postale dostupne tek razvojem odgovaraju?ih fizi?kih metoda. Najva?nija od njih bila je difrakcija rendgenskih zraka. Iz obrazaca raspr?enja rendgenskih zraka na kristalima postalo je mogu?e odrediti ta?an polo?aj atoma u kristalu, a za molekularne kristale bilo je mogu?e lokalizirati atome u jednoj molekuli. Druge metode uklju?uju difrakciju elektrona dok prolaze kroz plinove ili pare i analizu rotacijskih spektra molekula.
Sve ove informacije daju samo op?u ideju o strukturi molekula. Priroda hemijskih veza mo?e se istra?iti modernom kvantnom teorijom. I iako se molekularna struktura jo? ne mo?e izra?unati s dovoljno visokom precizno??u, svi poznati podaci o kemijskim vezama mogu se objasniti. ?ak je i predvi?eno postojanje novih vrsta hemijskih veza.
Jednostavna kovalentna veza. Molekul vodonika H 2 sastoji se od dva identi?na atoma. Prema fizi?kim mjerenjima, du?ina veze - udaljenost izme?u jezgara atoma vodika (protona) - je 0,70(1 = 10 -8 cm), ?to odgovara polupre?niku atoma vodika u osnovnom stanju, tj. u najni?em energetskom stanju. Veza izme?u atoma mo?e se objasniti samo pretpostavkom da su njihovi elektroni lokalizirani uglavnom izme?u jezgara, formiraju?i oblak negativno nabijenih vezivnih ?estica i dr?e?i pozitivno nabijene protone zajedno.Razmotrimo dva atoma vodika u osnovnom stanju, tj. stanje u kojem su njihovi elektroni na 1
s -orbitale. Svaki od ovih elektrona se mo?e zamisliti kao talas, a orbitala kao staja?i talas. Kada se atomi pribli?e jedan drugom, orbitale po?inju da se preklapaju (slika 2), i, kao iu slu?aju obi?nih talasa, dolazi do interferencije – superpozicije talasa (talasnih funkcija) u oblasti preklapanja. Ako su predznaci valnih funkcija suprotni, tada se valovi me?usobno uni?tavaju tijekom interferencije (destruktivna interferencija), a ako su isti, onda se dodaju (konstruktivna interferencija). Kada se atomi vodika pribli?e jedan drugom, mogu?a su dva ishoda u zavisnosti od toga da li su valne funkcije u fazi (slika 2,a ) ili u antifazi (slika 2,b ). U prvom slu?aju do?i ?e do konstruktivne interferencije, u drugom do destruktivne interferencije i pojavit ?e se dvije molekularne orbitale; jedan od njih karakteri?e velika gustina u podru?ju izme?u jezgara (slika 2,in ), za drugi - nizak (sl. 2,G ) je zapravo ?vor sa nultom amplitudom koji razdvaja jezgra.Dakle, kada se atomi vodika me?usobno pribli?avaju i njihova interakcija 1
s -orbitale formiraju dvije molekularne orbitale, a dva elektrona moraju ispuniti jednu od njih. Elektroni u atomima uvijek nastoje zauzeti najstabilniji polo?aj – onaj u kojem je njihova energija minimalna. Za orbitalu prikazanu na sl. 2,in , velika gustina je karakteristi?na u podru?ju izme?u jezgara, a svaki elektron koji zauzima ovu orbitalu bit ?e bve?inu vremena biti u blizini pozitivno nabijenih jezgara, tj. njegova potencijalna energija ?e biti mala. Naprotiv, orbitala prikazana na Sl. 2,G , maksimalna gusto?a se odvija u podru?jima koja se nalaze lijevo i desno od jezgara, a energija elektrona smje?tenih u ovoj orbitali bit ?e velika. Dakle, elektroni imaju manju energiju kada zauzmu orbituin , a ova energija je ?ak i manja od one koju bi imali da su atomi beskona?no udaljeni jedan od drugog. Po?to u ovom slu?aju postoje samo dva elektrona, oba mogu zauzeti energetski povoljniju orbitu ako su njihovi spinovi antiparalelni (Paulijev princip). Dakle, energija sistema koji se sastoji od dva atoma vodika opada kako se atomi pribli?avaju jedan drugom, a da bi se potom atomi uklonili jedan od drugog, energija jednaka energiji formiranja stabilne molekule vodika H 2 . Imajte na umu da je neophodan uslov za postojanje molekule vodika preferencijalna lokalizacija elektrona izme?u jezgara u skladu sa onim ?to smo ve? rekli gore. molekularna orbitalain naziva se orbitala vezivanja i orbitalaG - olabavljenje.Razmotrimo sada pristup dva atoma helijuma (atomski broj 2). I ovdje se preklapa 1
s -orbitale dovodi do stvaranja dvije molekularne orbitale, od kojih jedna odgovara ni?oj, a drugoj vi?oj energiji. Ovaj put, me?utim, potrebno je postaviti 4 elektrona u orbitale, po 2 elektrona iz svakog atoma helija. Samo dva od njih mogu ispuniti niskoenergetsku veznu orbitalu, druga dva moraju zauzeti orbitalu visoke energijeG . Smanjenje energije zbog povoljnog polo?aja prvog para pribli?no je jednako pove?anju energije zbog nepovoljnog polo?aja drugog para. Sada pristup atoma ne daje dobit u energiji, a molekularni helijum ne 2 nije formirana. Ovo se mo?e zgodno ilustrovati dijagramom (slika 3); razli?ite orbitale na njemu su predstavljene u obliku energetskih nivoa na kojima se mogu locirati elektroni. Potonje su ozna?ene strelicama koje pokazuju gore i dolje kako bi se razlikovali smjerovi okretanja. Dva elektrona mogu zauzeti istu orbitalu samo ako su njihovi spinovi antiparalelni.Ovi op?i principi su ispunjeni u formiranju molekula iz atoma. ?im se dva atoma pribli?e tako blizu da se njihove atomske orbitale (AO) po?nu preklapati, pojavljuju se dvije molekularne orbitale (MO): jedna se vezuje, druga labavi. Ako svaki od AO ima samo jedan elektron, oba mogu zauzeti vezni MO sa ni?om energijom od AO i formirati hemijsku vezu. Veze ovog tipa, koje se danas nazivaju kovalentne, odavno su poznate kemi?arima (ideja o kovalentnoj vezi bila je osnova oktetne teorije veze koju je formulirao ameri?ki fizikalni hemi?ar G. Lewis 1916. godine). Njihovo formiranje obja?njeno je socijalizacijom para elektrona me?usobnom interakcijom atoma. Prema modernim konceptima, snaga veze zavisi od stepena preklapanja odgovaraju?ih orbitala. Sve navedeno sugerira da se veze izme?u atoma mogu formirati tokom socijalizacije ne samo dva, ve? i jednog ili tri elektrona. Me?utim, one ?e biti slabije od konvencionalnih kovalentnih veza iz sljede?ih razloga. Kada se formira jednoelektronska veza, energija samo jednog elektrona opada, a u slu?aju stvaranja veze kao rezultat socijalizacije tri elektrona, energija dva od njih opada, dok tre?eg, naprotiv, raste, kompenzuju?i smanjenje energije jednog od prva dva elektrona. Kao rezultat toga, nastala veza od tri elektrona je dvostruko slabija od uobi?ajene kovalentne veze.
Socijalizacija jednog i tri elektrona nastaje tokom formiranja molekularnog vodonikovog jona H
2 + i HHe molekule, respektivno. Op?enito, veze ovog tipa su rijetke, a odgovaraju?i molekuli su visoko reaktivni.Ve?ina supstanci koje stvaraju film su oligomeri i polimeri.
Oligomeri su polimeri male molekularne te?ine (obi?no ne vi?e od nekoliko hiljada). Oligomeri se ?esto nazivaju polimeri sa stepenom polimerizacije manjim od onog pri kojem se po?inju manifestirati specifi?na svojstva polimera povezana s fleksibilno??u njegovih makromolekula. Od velikog zna?aja su oligomeri koji sadr?e funkcionalne grupe koje odre?uju sposobnost molekula oligomera da se me?usobno kombinuju i formiraju duge molekularne lance ili trodimenzionalne mre?ne strukture (fenol-formaldehidne smole u fazi rezole, epoksidne smole, poliester akrilati, koji polimerizuju zbog terminalnih dvostrukih veza i sl.).
Polimeri su makromolekularna jedinjenja ?ije se makromolekule sastoje od velikog broja ponavljaju?ih jedinica ?ija se molekulska te?ina kre?e od nekoliko hiljada do nekoliko miliona. Molekuli makromolekularnih jedinjenja (makromolekule) uklju?uju stotine i hiljade atoma me?usobno povezanih silama glavnih valencija.
Govore?i o strukturnoj formuli makromolekula, napominjemo da atomi ili atomske grupe u molekuli makromolekularnog spoja mogu biti raspore?eni ili u obliku dugog lanca (linearnog, na primjer, celuloze), ili u obliku dugog lanca sa granama (razgranatim, na primjer, amilopektin), ili, kona?no, u obliku trodimenzionalne mre?e koja se sastoji od segmenata lan?ane strukture (pro?ivene). Primjer umre?enih spojeva visoke molekularne te?ine su fenolne smole.
Ako se molekularni lanci makromolekula sastoje od velikog broja ponavljaju?ih grupa - jedinica koje imaju istu strukturu, onda se takva visokomolekularna jedinjenja nazivaju polimeri; ako sadr?e vi?e vrsta ponavljaju?ih grupa - kopolimeri - A - B - A - B -.
U zavisnosti od hemijskog sastava glavnog lanca, makromolekularna jedinjenja se dele na heterolan?ane, u glavnom lancu makromolekula koji sadr?e atome razli?itih elemenata (ugljik, azot, silicijum, fosfor), i homolan?ane, ?iji su makromolekularni lanci gra?eni od identi?nih atoma, glavno mjesto me?u njima zauzimaju karbolan?ani polimeri (glavni lanci makromolekula sastoje se samo od atoma ugljika). Ako polimerne makromolekule, zajedno s atomima ugljika, sadr?e atome neorganskih elemenata, onda se nazivaju organoelementima.
Hemijska struktura nekih predstavnika polimera izgleda ovako:
… -
CH2 - CH2 - CH2 - CH2 - ...
fragment polimera ugljeni?nog lanca (polietilen, polipropilen, poliizobutilen, polimetil metakrilat, polivinil alkohol, itd.).
Polimeri iz klase heterolanaca uklju?uju brojne jednostavne i slo?ene poliestere, poliamide, poliuretane, prirodne proteine itd., kao i veliku grupu polimera organskih elemenata:
… -
CH2 - CH2 - O - CH2 - CH2 - O - CH2 - CH2 - O -
polietilen oksid (eter)
polietilen tereftalat (poliester)
poliamid
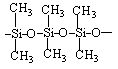
polidimetilsiloksan (polimer organskih elemenata)

polifosfonitril hlorid (anorganski polimer).
Druga klasa polimera su visokomolekularna jedinjenja sa sistemom konjugovanih veza: razli?iti poliacetileni, polinitrili, polifenileni, polioksadiazoli i mnoga druga jedinjenja. Na primjer,
... - CH = CH - CH = CH - CH = CH - ...
poliacetilen
polifenilen
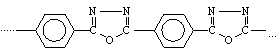
polioksadiazol
 Ova klasa tako?er uklju?uje grupu helatnih polimera, koji uklju?uju razli?ite elemente koji su sposobni za formiranje koordinacionih veza (obi?no su ozna?eni strelicama). Osnovna jedinica takvih polimera ?esto ima slo?enu strukturu:
Ova klasa tako?er uklju?uje grupu helatnih polimera, koji uklju?uju razli?ite elemente koji su sposobni za formiranje koordinacionih veza (obi?no su ozna?eni strelicama). Osnovna jedinica takvih polimera ?esto ima slo?enu strukturu:
Svojstva helatnih polimera jo? nisu dovoljno prou?ena, ali su, nesumnjivo, vrlo interesantna sa vi?e stajali?ta.
U zavisnosti od oblika makromolekula, makromolekularna jedinjenja se dele na fibrilarna i globularna. U fibrilarnim polimerima, molekule su linearnog ili blago razgranatog lanca. Fibrilarni makromolekularni spojevi lako formiraju supramolekularne strukture u obliku asimetri?nih paketa molekula - fibrila. Lanci molekula unutar svake fibrile su orijentisani u istom pravcu (celulozna vlakna, poliamidi, itd.).
Visokomolekularna jedinjenja nazivaju se globularna, ?ije makromolekule imaju oblik vi?e ili manje sfernih zavojnica, globula, potonje mogu biti visoko razgranate makromolekule. Uni?tenje takve globule nemogu?e je bez hemijskog uni?tenja makromolekula. Tako?er je mogu?e formiranje globula u fibrilarnim makromolekularnim spojevima povezano s promjenom oblika njihovih molekula.
Pojedina?na globula mo?e biti formirana od fleksibilne linearne makromolekule smotane u kuglu pod utjecajem sila unutarmolekulske interakcije.
Struktura polimera u obliku dugih makromolekula, ?ije su karike povezane hemijskim vezama du? lanca makromolekula, dokazana je na razli?ite na?ine. Jedna od mo?nih metoda za analizu hemijske strukture molekula je metoda analize rendgenske difrakcije. Pomo?u ove metode mogu?e je odrediti udaljenost izme?u centara mase susjednih atoma povezanih kako kemijski tako i intermolekularnim (fizi?kim) vezama. Budu?i da su du?ine hemijskih veza u kristalnim supstancama male molekulske mase dobro poznate, mogu?e je analizom rendgenske difrakcije orijentisanih polimernih sistema u kristalnom stanju odrediti koji se od atoma nalazi na udaljenosti od svakog od njih. drugi jednaki du?ini hemijske veze, a koji su na ve?oj udaljenosti, odnosno hemijski nepovezani. Na ovaj na?in je utvr?eno prisustvo hemijskih veza du? lanca makromolekula i me?umolekulskih veza susednih atoma kako unutar lanca tako i izme?u susednih lanaca.
Treba napomenuti da prisustvo mnogih jedinica u makromolekuli odre?uje raznolikost hemijske strukture polimera. Na primjer, svaka karika u procesu elementarnog ?ina rasta lanca mo?e biti pri?vr??ena za susjednu kariku na razli?ite na?ine - "glava na glavu", "rep na rep" ili "glava na rep". Mogu?e su razli?ite opcije za pri?vr??ivanje veze na rastu?u makromolekulu za asimetri?ne monomere tipa
Za takve monomere mogu?e su direktne opcije:

i "od glave do repa":

Tako?er je mogu?e mijenjati tipove povezivanja. Dakle, polimer ne sadr?i makromolekule potpuno iste kemijske strukture, ve? mje?avinu izomernih makromolekula, ?to ga razlikuje od ?istih tvari male molekularne mase izgra?ene od identi?nih molekula.
U nizu supstituiranih zasi?enih ugljikovodika, s pove?anjem broja atoma ugljika, broj mogu?ih izomera brzo raste. A kada broj veza dosegne desetine ili stotine hiljada, tada ?e broj mogu?ih izomera biti izra?en astronomskim brojevima.
Polimeri izgra?eni od jedinica s pravilno izmjenjivim smjerovima supstituenata nazivaju se stereoregularni.

Ako su supstituenti locirani na jednoj strani ravnine glavnih veza, stereoregularni polimeri se nazivaju izotakti?kim, ako su s obje strane - sindiotakti?kim.
Ako su supstituenti nasumi?no raspore?eni na obje strane ravnine glavne veze, nazivaju se nepravilnim ili atakti?kim:
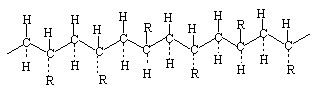
Struktura polimera dobivenih iz disupstituiranih monomera je jo? slo?enija, jer se ve? u samom monomeru supstituenti mogu nalaziti na jednoj strani primarne veze (cis-izomer) ili na obje strane (trans-izomer):
cis-trans-
Sinteza makromolekula iz cis-izomera dovodi do proizvodnje eritrodiizotakti?kih polimera:
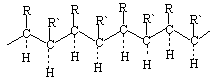
i od trans-izomera - treodiizotakti?kih polimera:
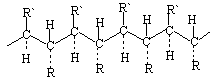
Sve promjene u kemijskoj strukturi polimera podrazumijevaju promjenu svojstava materijala na njihovoj osnovi.
Pitanja vezana za stereoregularnost gra?enja makromolekularnih lanaca su vrlo interesantna, ali ni?ta manje slo?ena i privla?e pa?nju istra?iva?a ve? dugi niz godina. Materijali na bazi stereoregularnih polimera imaju vrlo razli?ita svojstva od onih na bazi nepravilnih polimera. Lako se kristaliziraju, osiguravaju?i regulaciju njihove fizi?ke strukture i svojstava, kao i ?irenje temperaturnih granica performansi. Klasi?an primjer "popre?no povezanih" polimera su o?vr??ene epoksidne smole:
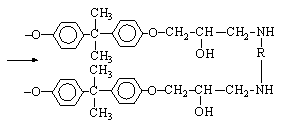
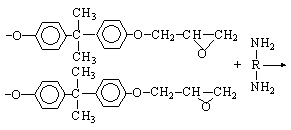
Ako su svi glavni lanci u bloku polimernog materijala me?usobno povezani prostornim vezama ili lancima, blok se mo?e smatrati jednom divovskom makromolekulom.
Sada je utvr?eno da strukturirani polimeri kao ?to su fenol-formaldehidne i epoksidne smole uop?e ne tvore pravilnu prostornu mre?u sa sekvencijalnom izmjenom veza i popre?nih veza. Blok takvog prostorno strukturiranog polimera ?esto se gradi od globula nastalih ve? u procesu sinteze smole. Globularne formacije koje se pojavljuju u prvoj fazi sinteze otapaju monomer, a daljnji proces kondenzacije sastoji se u pove?anju veli?ine globula. U zavr?noj fazi procesa, globule sadr?e veliki broj makromolekula. Ovo ne ometa naknadno hemijsko povezivanje izme?u pojedina?nih niti.
Postoji nekoliko glavnih na?ina za dobivanje mre?nih polimera:
1. Izvo?enje hemijske reakcije izme?u dvije (ili vi?e) razli?itih funkcionalnih krajnjih grupa vezanih za lanac male molekularne te?ine. Kao rezultat, formira se gusta mre?a s kratkim lancima izme?u popre?nih veza.
2. Hemijsko vezivanje jedinjenja visoke molekularne te?ine krajnjim grupama kori?tenjem agensa za umre?avanje niske molekularne te?ine. Kao rezultat, formira se rijetka mre?a s pro?irenim linearnim fragmentima izme?u ?vorova ?ava.
3. Formiranje mre?e zbog kopolimerizacije dvo- i polifunkcionalnih monomera. Primjer takve mre?e je sistem stiren - divinilbenzen:

4. Vulkanizacija polimernih lanaca uklju?ivanjem funkcionalnih grupa koje se nalaze du? glavnog lanca u reakciju. Reakcija se provodi ili kori?tenjem agensa za umre?avanje niske molekularne te?ine, ili zbog zra?enja i drugih vrsta izlaganja funkcionalnim grupama.
5. Formiranje mre?a zbog reakcije dva (ili vi?e) razli?ita polimera na funkcionalnim grupama koje se nalaze du? lanca svakog od polimera (tj. u jedinicama koje se ponavljaju, a ne na krajevima).
6. Sinteza polimernih mre?a reakcijom policiklotrimerizacije. Za to se koriste oligomeri sa krajnjim grupama koje mogu formirati cikluse tokom reakcije. Na primjer, trimerizacija bifunkcionalnih oligomera (ili monomera) koji sadr?e cijanatne krajnje grupe:
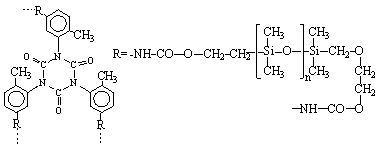
Mogu?i su i drugi na?ini dobijanja polimernih mre?a.
Relativno nova vrsta polimera su "interpolimeri", pod kojima se podrazumeva sistem izgra?en od dva (ili vi?e) hemijski heterogenih makromolekula, hemijski povezanih jedna s drugom zbog funkcionalnih grupa sme?tenih u ponavljaju?im jedinicama svakog makromolekula. Ovo je ?ematski prikazano na Sl. jedan.
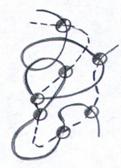
Rice. 1. ?ematski prikaz interpolimerne makromolekule
Specifi?an primjer takvog sistema je proizvod interakcije polistirena sa politriklorobutadienom:

Priprema interpolimera omogu?ava otvaranje mogu?nosti za modifikaciju strukture i svojstava polimera.
Dakle, prema modernim idejama o strukturi i svojstvima polimera, struktura polimera po?inje ve? oblikom makromolekule i njenom lokacijom u prostoru. Makromolekula je uvijek primarni element bilo koje strukture (ba? kao ?to je elementarna karika primarni element kemijske strukture lanca).
U tehnologiji boja i lakova, hemijski procesi koji dovode do stvaranja prostornih polimera zauzimaju posebno mesto i va?na su tehnika za dobijanje premaza sa ?eljenim svojstvima. Dakle, alkidni, fenol- i amino-formaldehidni, epoksidni, poliuretanski i drugi tvorci filma u smislu molekularne te?ine i svojstava prolaze kroz hemijsko o?vr??avanje tokom formiranja filma.
Prelazak filmotvoraca u stanje "mre?nog" polimera prostorne strukture dovodi do pobolj?anja atmosferske, vodene, uljne i toplinske otpornosti, tvrdo?e i ?vrsto?e, prianjanja itd.
Svi termoreaktivni tvornici filma sadr?e reaktivne grupe, ali u nekim slu?ajevima je potreban dodatni reagens za formiranje prostornog polimera - u?vr??iva?a, kao da "popre?no povezuje" makromolekule jedni s drugima.
Prekomjerna u?estalost me?umolekularnih veza dovodi do pove?anja krhkosti i sklonosti filma pucanju zbog pove?anja unutra?njih naprezanja. Zbog toga se u?estalost veza regulira promjenom na?ina o?vr??avanja (temperatura, trajanje), broja funkcionalnih grupa formira?a filma, kao i prirode i sadr?aja o?vr??iva?a.
Razmotrimo prvo dva objekta - dijamant i kalcit, ?ija je struktura prili?no tipi?na za obi?nu materiju:
Supstance ove vrste se ?esto nalaze u prirodi. Vidimo da imaju ure?enu formu, a za to postoje razlozi, koji ?e postati jasni kada se supstanca podijeli na sve manje dijelove. Ostavimo dijamant po strani (na? bud?et nam ne dozvoljava da eksperimenti?emo s njim) i po?nemo drobiti kalcit dlijetom i ?eki?em. Raspa??e se na male komadi?e, ali - ?to je najzanimljivije - ovi komadi ?e ponoviti strukturu velikog komada. Zanemaruju?i dimenzije, mo?ete vidjeti da uglovi izme?u lica i ravni ostaju konstantni. Zdrobiv?i mineral na najsitnije ?estice i ispitaju?i ih pod mikroskopom, vidjet ?emo isti oblik koji nam je ve? poznat. Ispostavilo se da ?ak i najmanje ?estice materije imaju takvu strukturu.
Hemi?ari koji kalcit nazivaju kalcij karbonatom re?i ?e da se njegova struktura sastoji od karbonatne grupe (CO 3 u kojoj je atom ugljika vezan za tri atoma kisika) i jednog atoma kalcija. Fizi?ka zapa?anja pokazuju da se brojne karbonatne grupe i atomi kalcija nalaze u prostoru pod istim uglovima kao i lica velikog kristala kalcita.
Dakle, vidljiva struktura materijala ponavlja kristalnu strukturu. Ovo je ista struktura, samo mnogo puta ve?a.

Fizi?ka svojstva materije na makroskopskom nivou odra?avaju obrasce na mikroskopskom nivou.
Strukturu biolo?kog materijala odre?uje i njegova molekularna struktura. Mnoge biolo?ke strukture izgledaju kao kristali, a pod mikroskopom mo?ete vidjeti njihove lijepe, jasne oblike. Ve? smo vidjeli kako su ?elije unutar organizma ure?ene. Ovaj raspored ovisi o strukturi materijala od kojih su sastavljeni.
?elije i tkiva svih organizama sastoje se od istih supstanci. Prije svega, ovo vode. Voda ?ini oko 70-90% svih biolo?kih supstanci, te stoga fizi?ka i hemijska svojstva vode u velikoj mjeri odre?uju svojstva biolo?kog materijala. U vodi se otapaju soli elemenata kao ?to su natrijum, kalijum, kalcijum, magnezijum i hlor. Preostali dio je za organska materija, koji se sastoje od atoma ugljika (C) vezanih za vodonik, kisik, du?ik (N), a ponekad i atome sumpora (S) i fosfora (P).
Najjednostavniji organski molekuli koji se nalaze u prirodnom plinu ili nafti su metan, etan i propan.

Zovu se ugljikovodici jer se sastoje od atoma ugljika i vodika. Ovi atomi se mogu prikazati kao male kuglice povezane jedna s drugom. hemijske veze. U hemijskoj vezi, dva atoma dijele par elektrona, po jedan iz svakog atoma. Na na?im crte?ima, veza izme?u dva atoma je prikazana kao linija. Svaki element je karakteriziran valencija, ili sposobnost formiranja odre?enog broja hemijskih veza. Valencija ugljika je ?etiri, tako da svaki atom ugljika mo?e biti vezan za ?etiri druga atoma; zbog ovog svojstva nastaje veliki broj vrlo razli?itih kombinacija atoma, ?to dovodi do ogromne raznolikosti organskih molekula (slika 3.3). Dvije i tri paralelne linije zna?e dvostruku, odnosno trostruku vezu. Vezivanje kroz par elektrona naziva se kovalentna; veoma je jak, potrebna je zna?ajna koli?ina energije da se razbije, jer su organski molekuli prili?no stabilni. Me?utim, veze se prilikom sagorijevanja (oksidacije) lako raskidaju, osloba?aju?i veliku koli?inu energije, pa ugljovodonici slu?e kao vrijedno gorivo.
U najjednostavnijoj organskoj molekuli metana, atom ugljika je vezan za samo ?etiri atoma vodika. U drugoj molekuli, atom ugljika je povezan jednom vezom s drugim atomom ugljika, formiraju?i C-C lanac, na ?ijim se krajevima nalaze atomi vodika. C-C lanac mo?e biti veoma duga?ak; molekule voska, na primjer, sastoje se od 30-36 atoma ugljika. Lanac ugljikovih atoma tako?er mo?e biti zatvoren u prstenove razli?itih veli?ina. Ali najve?a raznolikost dolazi od kombinacije atoma ugljika sa grupama atoma drugih elemenata. Na primjer, OH (kiseonik vezan za vodonik) hidroksilna grupa vezana za ugljikov lanac formira se alkohol (alkohol).

Rice. 3.3. Razli?ite organske molekule, ?iji su glavni element atomi ugljika, obi?no povezani u lance. Svaka linija izme?u atoma odgovara vezi, odnosno zajedni?kom paru elektrona. Dvostruke i trostruke linije predstavljaju dvostruke i trostruke veze izme?u atoma. Slo?eniji molekuli, posebno oni s prstenastom strukturom, obi?no se prikazuju kao linije, gdje atomi ugljika (?esto s jednim ili dva atoma vodika) nisu nazna?eni na spojevima. Po?to je valencija ugljenika ?etiri, svaki ugljenik mora imati ?etiri veze; ako su prikazane samo tri veze atoma ugljika, tada se na ovaj atom mora vezati jo? jedan atom vodika
Amino grupa, koja se sastoji od atoma du?ika i dva atoma vodika (NH 2), povezana s ugljikovim lancem, formira amin. U slo?enijim grupama, atom kisika je vezan za atom ugljika dvostrukom vezom (C=O), a jedna od ovih kombinacija, karboksilnu grupu COOH formira molekul kiseline. (Kiselina je bilo koje hemijsko jedinjenje koje se formira joni vodonik; Podsjetimo da se pozitivno i negativno nabijeni atomi ili grupe atoma nazivaju ioni.)
Kombinacije svih vrsta ovih grupa sa uglji?nim lancima razli?itih du?ina i prstenova daju neobi?no veliki broj organskih spojeva, ali se samo nekoliko njih ?esto nalazi u ?ivim organizmima. Najva?niji spojevi su proteini, nukleinske kiseline, ugljikohidrati i lipidi.
lipidi, koji uklju?uju dobro poznate masti i ulja, sastoje se od dugih ugljikovih lanaca - obi?no od 16-18 atoma ugljika. Dobro smo svjesni njihovih svojstava: na kraju krajeva, upravo to su tvari koje ostavljaju neizbrisive mrlje na odje?i. Svi znaju da se voda i ulje ne mije?aju. Tvari koje se mije?aju s vodom nazivaju se hidrofilna(bukvalno, "vola koja voli"), i tvari koje se, poput ulja, ne mije?aju s njom, nazivaju se hidrofobna(“strah od vode”). (Masne, masne mrlje na odje?i treba ukloniti sredstvima za kemijsko ?i??enje koja sadr?e rastvara?e kao ?to je tetrahlorid ugljikohidrata, ili rastvara?ima koji sadr?e benzin, koji je tako?er hidrofoban.) U su?tini, lipidi se mogu definirati kao tvari koje se otapaju samo u hidrofobnim rastvara?ima.
Druge va?ne biolo?ke supstance odlikuju se gigantskom veli?inom svojih molekula. Molekularna te?ina malih molekula, kao ?to su propan, benzin ili ?e?er (poput glukoze), ne prelazi dvije stotine jedinica. Za razliku od njih, proteine, nukleinske kiseline i neke druge gra?evne materijale ?elija formiraju velike molekule - makromolekule, jer se njihova molekularna te?ina kre?e u hiljadama i vi?e. Nije neuobi?ajeno da gra?evinski materijali kaveza mogu biti toliki, jer u konstrukciji koristimo i duga?ke ?eli?ne grede i stropove od ?perplo?e i armiranog betona. ?vrsti dijelovi ?elija tako?er se sastoje od velikih komponenti.
Ali sve ove makromolekule imaju relativno jednostavnu strukturu. Oni predstavljaju polimeri, koji se sastoji od ponavljanja istih ili identi?nih molekula, tzv monomeri:

Na primjer, ugljikovodici se sastoje od ?e?era, koji su male organske molekule s formulom poput C 6 H 12 O 6 . ?e?eri od najve?eg interesa za nas – kao ?to su glukoza, galaktoza i manoza – imaju slo?enu strukturu. Mogu se povezati jedni s drugima, formiraju?i duga?ke lance, ponekad ?ak i sa granama. Kada se molekuli glukoze pove?u na specifi?an na?in (hemi?ari to zovu beta 1:4 veza), dobijaju celuloza:
Celuloza je jak vlaknasti materijal koji ?ini zidove biljnih ?elija, te je kao rezultat glavna komponenta drveta. Ali ako se molekule glukoze kombiniraju druga?ije (veza alfa 1:4, ponekad s ograncima 1:6), tada se dobivaju ?krob i glikogen - glavni rezervni materijal biljaka i ?ivotinja. Ostali ?e?eri u raznim jedinjenjima formiraju pektine i gume, koji ?ine so?nu pulpu vo?a i drugih dijelova biljaka. Svi ovi polimeri, ?ija masa dose?e nekoliko hiljada jedinica, nazivaju se polisaharidi, i njihovi sastavni monomeri (?e?eri) - monosaharidi. Drugi polimeri tako?e imaju imena koja po?inju prefiksom "poli", ?to zna?i "puno".
Jedan od najva?nijih polimera proteini, sastoje se od dugih lanaca monomera amino kiseline. Aminokiseline su tako nazvane jer sadr?e amino grupu (NH 2) i grupu organske kiseline (COOH). Dvije aminokiseline su povezane povezivanjem karboksilne grupe jedne s amino grupom druge i osloba?anjem molekule vode:

Rezultiraju?a molekula (dipeptid) jo? uvijek ima amino grupu na jednom kraju i kiselu grupu na drugom, tako da se druge aminokiseline mogu vezati za njega. Nastaju tri aminokiseline tripeptid, i tako dalje; zove se molekul mnogih aminokiselina polipeptid koji je, u stvari, protein. U tipi?nom proteinu, 200-300 aminokiselina je povezano u jedan dugi lanac. (Kada aminokiselina izgubi amino grupu i kiselinsku grupu, integri?u?i se u lanac, naziva se ostatak aminokiseline.) Po?to prosje?na aminokiselina ima molekulsku te?inu od oko 100 jedinica, lanac od 300 aminokiselina, ili prosje?ni protein, ima atomsku te?inu od oko 3000 jedinica.
Prirodni proteini se formiraju od 20 vrsta aminokiselina koje se razlikuju samo po strukturi bo?nog lanca (tabela 3.1). Aminokiseline se mogu kombinovati bilo kojim redoslijedom, tako da stanice mogu proizvesti ogroman broj vrsta proteina. Njihova navodna raznolikost je izvan ljudske koncepcije. Ako postoji 20 vrsta aminokiselina, onda 2 aminokiseline - 400 vrsta dipeptida (sa dva ostatka). Ve? ?e postojati 8.000 tripeptida, 160.000 tetrapeptida i 20.300 vrsta lanaca od 300 aminokiselina. Tako ogroman broj je nezamisliv. Svi proteini koje su ikada proizveli kopneni organizmi samo su mali dio mogu?e raznolikosti.
Svaka vrsta proteina ima jedinstvenu sekvencu aminokiselina. Na primjer, kod ljudi, molekula hemoglobina, koja je dio crvenih krvnih zrnaca - eritrocita, prenosi kisik krvlju. Po?inje sekvencom Val-H is-Leu-Thr-Pro-Glu-Glu-Lys-Ser-Ala-Val-Thr-Ala (slovne skra?enice ozna?avaju jednu ili drugu aminokiselinu). Kod obi?nog ?ovjeka svaki molekul hemoglobina po?inje ovim nizom.

U najjednostavnijem organizmu proizvodi se najmanje oko 2 hiljade razli?itih proteina, a u slo?enim organizmima, na primjer, kod ljudi, oko 30-50 hiljada. (Nedavne studije su identifikovale upravo takav raspon, iako ta?an broj ostaje nepoznat.) Svaki protein ima strukturu pogodnu za razli?ite funkcije, budu?i da su proteini glavni "radni konji" tela. Oni obavljaju gotovo sve funkcije koje identificiramo s konceptom "?ivog organizma":
? proteini su enzimi, koji ubrzavaju i kontroli?u sve hemijske reakcije u telu;
? proteini formiraju vidljive strukture tijela: keratini slu?e kao gradivni blokovi kose, ko?e i perja; kolageni su dio hrskavice i kostiju;
? proteini formiraju vlakna koja ste?u i raste?u mi?i?e i druge pokretne strukture kao ?to su cilije i bi?evi;
? proteini ?ine va?nu klasu hormoni koji prenose signale od jedne vrste ?elija u telu do druge vrste ?elije;
? nastaju proteini receptori koji primaju signale povezuju?i se s drugim molekulima; ?elija prima signale od hormona ako se molekul hormona ve?e za jedan od njegovih
receptori; receptori, zahvaljuju?i kojima osje?amo okus i miris, omogu?avaju tijelu da prepozna prisustvo malih molekula u vanjskom okru?enju i odgovori na njih; proteini prenose ione i male molekule kroz ?elijske membrane, ?to je neophodno za funkcionisanje na?eg nervnog sistema i sl.
organi poput bubrega; proteini reguli?u sve vrste procesa i prate
kako bi osigurali da se odvijaju odgovaraju?om brzinom.
Da biste razumjeli kako su ?elije raspore?ene i kako rade, mo?ete nau?iti samo vi?e o nekim funkcijama proteina.
Kraj rada -
Ova tema pripada:
Genetika
GENETIKA.. VODI? ZA PO?ETNIKE.. B GUTTMAN A GRIFFITS D SUZUKI I T CULLIS..
Ako vam je potreban dodatni materijal na ovu temu, ili niste prona?li ono ?to ste tra?ili, preporu?ujemo da koristite pretragu u na?oj bazi radova:
?ta ?emo sa primljenim materijalom:
Ako vam se ovaj materijal pokazao korisnim, mo?ete ga spremiti na svoju stranicu na dru?tvenim mre?ama:
| tweet |
Sve teme u ovoj sekciji:
Guttman B., Griffith E., Suzuki D., Cullis T
G97 Genetika / Barton Guttman, Anthony Griffith, David Suzuki, Tara Cullis. - Per. sa engleskog. O. Perfilieva. - M.: FAIR-PRESS, 2004. - 448 str.: ilustr. - (Nauka i ?ivot).
Genetika: pro?lost, sada?njost i budu?nost
“Za?to Jimmy ima crvenu kosu kao ?to je njegova mama, a njegov tata crnu?” "Za?to ljudi nemaju ?tence?" “A ako se konj o?eni kravom, ho?e li imati djece?” „Za?to je Meri takva
Potraga za redom i smislom
Mikrobiolog i geneti?ar Fran?ois Jacob jednom je primijetio da “ljudski mozak jednostavno mora prona?i neki red u svemiru”. Svako dijete odmah po ro?enju nema sistem, sa
Moderna slika nauke
Genetika je najva?nija oblast moderne biologije, a da biste je bolje razumeli, prvo morate razumeti ?ta je nauka u celini. Nauka je vrsta ljudske aktivnosti, va?na
Perspektive moderne genetike
S obzirom na sociokulturni kontekst, razumljivo je za?to genetika izaziva takav interes i za?to otkri?a u njenom polju imaju tako dalekose?ne implikacije. Posljednjih godina, mj
Primitivno interesovanje za nasledstvo
Gledaju?i unatrag, dokazi o interesovanju za naslije?e mogu se na?i jo? u periodu paleolita, kada su ljudi tek po?eli da shvataju ?ta je reprodukcija. Uzmimo pirina? kao primjer.
Pripitomljavanje biljaka i ?ivotinja u ogledalu mita
U brojnim crte?ima, likovnim djelima i mitovima stari ljudi su odra?avali izgled svake od kultiviranih biljaka i pripitomljenih ?ivotinja, ?to je imalo vrlo va?an utjecaj na
Nau?ne teorije naslije?a
Svi primje?uju sli?nost djece i njihovih roditelja. Drevni ljudi su vjerovali da sli?ni ljudi imaju zajedni?ke pretke i stoga su posebnu pa?nju posvetili srodstvu. Pored toga ?to su se porodi?ne veze dr?ale na okupu
Odakle dolaze bebe?
Ogromna uloga naslije?a za dru?tvo, ne samo sa fiziolo?ke, ve? i sa kulturolo?ke ta?ke gledi?ta, kao i interes za zdravo potomstvo, natjerali su ?ovje?anstvo da razmi?lja o tome kako
?elijska struktura
Ba? kao ?to je teleskop revolucionirao astronomiju, mikroskop je pomogao ljudima da shvate od ?ega se sastoje ?ivi organizmi. Mo?e se zamisliti kakvo se iznena?enje i ?u?enje ogledalo na licima
Rast i biosinteza
Jedno od najo?itijih svojstava ?ivog organizma je sposobnost rasta. Rast bilo kojeg organizma, kao ?to je ?ovjek, rezultat je dva procesa: rasta stanica i diobe stanica. Chelov
Enzimi
Traku za monta?u u fabrikama opslu?uju ljudi (iako ih sada sve vi?e zamjenjuju roboti). Ko odr?ava puteve metabolizma u tijelu? Kako nastaju hemijske reakcije koje se okre?u jedan metar
Sinteza polimera
Tijekom primarnih metaboli?kih procesa sintetiziraju se sve aminokiseline, ?e?eri, lipidi i drugi molekuli malih stanica, koji idu na formiranje takvih makromolekula kao ?to su proteini i polisaharidi. P
?elije kao fabrike za samoreprodukciju i samoobnavljanje
Poku?ajmo ponovo zamisliti kako tijelo funkcionira. Iz okoline prima supstancu-sirovinu i raznim metaboli?kim putevima pretvara je u molekule svoje strukture – ?ini je od primarne
Revolucionarno otkri?e: Mendelovi zakoni
Misterija prenosa osobina naslije?em oduvijek je privla?ila ljude. U 1. veku pne e. Stari rimski filozof Lukrecije primijetio je da djeca ponekad li?e na svoje djedove ili pradjedove. Vek kasnije P
Mendelova otkri?a
Gregor Mendel je bio prvi koji se pribli?io razotkrivanju drevne misterije. Bio je monah u samostanu Brunn (danas Brno, ?e?ka) i, pored podu?avanja, bavio se eksperimentima na kri?anju
Rodovnici
Pored brojanja broja biljaka i ?ivotinja sa odre?enim osobinama dobijenim slu?ajnim ukr?tanjem, korisno je istra?iti mehanizam naslje?a na primjeru rodovnika (ljudi ili ku?e).
Krvne grupe
Dobra lekcija iz genetike mo?e biti prou?avanje krvnih grupa kod ljudi. Krv se svrstava u jednu ili drugu grupu u zavisnosti od toga kako je u interakciji sa imunolo?kim sistemom, koji ?titi
Vi?estruki aleli i dominacija
Fenomeni kao ?to su nepotpuna dominacija i kodominacija dokazuju da interakcija alela jednog gena mo?e biti prili?no slo?ena. Kao ?to smo vidjeli, krvnu grupu odre?uju tri alela od jednog
Test krstovi
Organizmi s dominantnim fenotipom za odre?enu osobinu mogu biti homozigotni ili heterozigotni - AA ili Aa, ako koristite konvencije. Ponekad je va?no znati genotip.
Vjerovatno?a
Mendelov zakon cijepanja omogu?ava vam da predvidite vjerovatno?u naslje?ivanja odre?enih osobina. G. Mendel se mo?e nazvati osniva?em statisti?kih metoda u prou?avanju genetike, jer
Dva ili vi?e gena
Ove principe teorije vjerovatno?e va?no je imati na umu kada analiziramo rezultat ukr?tanja dva ili vi?e gena u isto vrijeme. G. Mendel je provodio eksperimente u kojima je posmatrao simultano
Mendelov prvi zakon i definicija o?instva
Na osnovu jednostavnog Mendelovog rezonovanja, moderni geneti?ari odre?uju prirodu naslje?ivanja i ispoljavanje odre?ene osobine u pedigreima. Osim toga, Mendelovi zakoni ponekad mogu pomo?i u definiranju
?elije i reprodukcija
Nakon ?to je ?elijska teorija Schleidena i Schwanna postala op?eprihva?ena, patolog Rudolf Virchow je dao svoj zna?ajan doprinos. On je sugerisao da se ne samo svi organizmi sastoje od ?elija, ve? svaki
Mitoza i ?elijski ciklus
Jedna ?elija raste i deli se na dve nove ?elije dok prolazi kroz ?elijski ciklus. Svrha takvog ciklusa je da proizvede dvije ili vi?e identi?nih ?elija, koje ?e nastaviti proces, nakon ?to dobiju od roditelja
Kariotip
Poznavaju?i mehanizam mitoze, mo?ete bolje vidjeti hromozome koji se slobodno kre?u tokom ovog procesa. Stavimo kap krvi u epruvetu sa hranljivom otopinom u kojoj se leu mo?e razmno?avati.
Mejoza i Mendelovi zakoni
Danas je op?te poznato da su geni locirani na hromozomima, mada ?emo u narednom odeljku poku?ati da doka?emo ovu tvrdnju. Nakon ?to smo razmotrili proces mejoze, sada u njemu mo?emo prona?i razloge za red
Lokacija gena
Glavni procesi koji se de?avaju tokom mejoze i mitoze prou?avani su do kraja 19. veka. Sada je poznato da je to prili?no slo?en mehanizam za distribuciju hromozoma me?u ?elijama k?eri, ali sve do po?etka 20.
polni hromozomi
Jo? u davna vremena ljudi su primijetili da se neke bolesti javljaju gotovo isklju?ivo kod mu?karaca, iako se prenose po maj?inoj liniji. Najpoznatiji primjer je hemofilija, odnosno nedovoljna
Nedisjunkcija hromozoma
Obi?no mu?karci i ?ene imaju dobro definiran fenotip, odre?en njihovim skupom hromozoma - XY ili XX. Ali ponekad se djeca ra?aju s neobi?nim brojem spolnih kromosoma, a to se doga?a kao posljedica
Geni i metaboli?ki poreme?aji
Ljudi su lo? "materijal" za prou?avanje zakona naslije?a, jer je od njih te?ko dobiti dovoljno pouzdane podatke, ali su napravljena prva zapa?anja o tome kako geni obavljaju svoju funkciju i
Geni i enzimi
Godine 1944. George Beadle i Edward Tatham potvrdili su ispravnost Harrodovih zaklju?aka koriste?i kalup za kruh Neurospora (ova svijetlonarand?asta plijesan ponekad se formira na ustajalom kruhu) kao primjer.
Proteini i informacije
Po?to geni kontroli?u proizvodnju i sintezu proteina, pogledajmo ponovo strukturu proteina. Kao ?to je re?eno u gl. 3, proteini su najraznovrsniji molekuli u tijelu. One su integral
Korekcija nasljednih poreme?aja
Dok je genetika ?inila svoje prve korake po?etkom 20. stolje?a, ideja pobolj?anja ljudske rase, ili eugenika, dobila je veliku pa?nju (vidi Poglavlje 15). Kada su ljudi nau?ili mehanizam naslje?ivanja
bakterije
Podsjetimo da se bakterije razlikuju od drugih organizama po tome ?to su prokarioti, odnosno nemaju jezgro okru?eno membranom, za razliku od eukariota, uklju?uju?i biljke i ?ivotinje, u ?ijim stanicama
Prvi koraci
Godine 1928. Frederick Griffith je otkrio da supstanca mrtvih ?elija jednog soja bakterija mo?e prenijeti svoje karakteristike na ?ive ?elije drugog soja. Na primjer, poznat je soj IIIS
bakteriofagi
Godine 1915. Englez Frederick Twort i Kana?anin Felix D"Erell nezavisno su otkrili bakteriofage koji uzrokuju infekcije me?u bakterijama. Sama ideja infekcija me?u bakterijama
Hershey Chase eksperiment
Znaju?i da su fagi otprilike pola DNK, a pola proteini, Alfred Hershey i Martha Chase odlu?ili su istra?iti funkcije ove dvije komponente ozna?avaju?i ih, odnosno uklju?ivanjem u njih.
DNK struktura
Podsjetimo da su glavni gradivni blokovi tijela polimeri. Nukleinske kiseline su tako?er polimeri, iako se po strukturi veoma razlikuju od proteina. Tako?e ih zovu
DNK model i genetika
Za razliku od Mendelovog rada, Watsonov i Crickov rad je odmah privukao pa?nju nau?ne zajednice jer je objasnio mehanizam naslje?a. Odmah je postalo jasno da je
Validacija modela
Prava nau?na vrijednost modela mjeri se ?injenicom da je mogu?e testirati sve zaklju?ke do kojih on vodi u praksi. Watson-Crick model nije samo uklju?io sve poznate ?injenice o DNK i ljudima
Distribucija gena
?injenica da su geni locirani na hromozomima izgleda da nije u skladu sa ?injenicom da ljudi imaju samo 23 para hromozoma, a opet hiljade razli?itih osobina kojima se hiljade osobina moraju poklapati.
Sa h/Y: 1 Sa H/Y: 1 sa h/Y: 9 sa H/Y.
Ispostavilo se da je 10% sinova, koje nazivamo rekombinantima, dobilo druga?iju kombinaciju gena, razli?itu od kombinacije njihovih majki. U profazi mejoze gradim homologne parove
Sa H/Y: 1 Sa h/Y: 1 sa H/Y: 9 sa h/Y.
Ovo je za o?ekivati: 90% po?etnog rasporeda alela i 10% rekombinacija. Odre?ivanje udaljenosti izme?u ljudskih gena je prili?no te?ko. U ve?ini organizama ukr?tajte ma?ku
A c/Y, 7 A C/Y, 8 a c/Y, 42 a C/Y.
Ukupno se dobije 15 (7 + 8) rekombinacija od sto, odnosno 15%. Stoga se gen A mo?e smjestiti na hromozomsku mapu 15 jedinica od gena C. Me?utim, tri gena se mogu locirati u nizu
Crossover unutar gena
Sve do sredine 1940-ih, nau?nici su vjerovali da su geni najvjerovatnije hromomeri, odnosno sitne grudvice du? hromozoma, zahvaljuju?i
Genetika faga
Max Delbr?ck je za svoje istra?ivanje odabrao fage jer su oni vrlo jednostavan biolo?ki sistem: sitne ?estice koje se mogu razmno?avati u drugim stanicama.
Fina struktura gena
Seymour Benzer je prou?avao finu strukturu gena koriste?i T4 fage, me?u kojima je uspio izolirati rijetke intragene rekombinante. Benzer se fokusirao na klasu mutanata r - rII.
Komplementacija i razgrani?enje gena
Eksperimenti mapiranja su pokazali da se rII regija sastoji od mnogo malih regija, ili mjesta, gdje se mogu pojaviti razli?ite mutacije. Ali takve karte daju samo ideju
?ta je gen!
Vratimo se definiciji gena. U klasi?noj genetici, rije? "gen" ozna?avala je jedinicu genetskog materijala, raspore?enu prema tri kriterija: po funkciji, mutaciji i rekombinaciji. U po?etku se pretpostavljalo
Restrikcijski enzimi i palindromi
Bakterije i fagi koji ih napadaju u stanju su kontinuiranog hemijskog rata. Bakterije koje se odupiru infekciji faga dobijaju prednost u borbi za egzistenciju, i one
Mapiranje ograni?enja
Mnoge vrste restrikcijskih enzima su sada poznate i dostupne za upotrebu. Oni seku DNK u razli?ite sekvence i mogu se koristiti za analizu strukture DNK i kompajliranje
Kako se grade proteini?
Dakle, informacija koja odre?uje redoslijed aminokiselina u proteinu pohranjena je u DNK u obliku niza triplet kodona. Ali kako se sekvenca DNK baza pretvara u pravi proizvod? Naravno, crte?i
RNA molekuli: alati za sintezu proteina
U 1940-im godinama, kada nau?nici jo? uvijek nisu imali dobro razumijevanje strukture nukleinskih kiselina, dobijeni su dokazi da je sinteza proteina uvijek pra?ena sintezom ribonukleinske kiseline.
RNA transkripcija
Sada je dokazano da RNK nastaje kao rezultat istog uparivanja komplementarnih baza, uz pomo? kojih se iz jednog lanca formira dvostruka spirala DNK (slika 9.2). Ovaj proces se zove
Broadcast
Prijenos informacija sa DNK na RNK naziva se transkripcija, a prijenos ovih informacija sa mRNA na protein se naziva translacija. Messenger RNA obi?no programiraju ribozom neko vrijeme.
Kompleksni eukariotski geni
Kada su istra?iva?i po?eli prou?avati gene razli?itih proteina u eukariotskim stanicama, otkrili su da je interakcija gena i proteina u ovim organizmima slo?enija od interakcije gena i proteina u proci.
Genetski rje?nik
Do 1962. godine, zahvaljuju?i ranije spomenutom radu Cricka i njegovih kolega, ustanovljeno je da se genetski kod sastoji od trojki. Nakon toga, istra?iva?i su se suo?ili sa jo? jednim te?kim problemom.
Kolinearnost gena i proteina
Hipoteza kolinearnosti gen-protein mogla bi se potvrditi pokazivanjem da sekvenca genskih mutacija odgovara promjenama u sekvenci aminokiselina do kojih te mutacije dovode. Za
Terminacijski kodoni
Tri ona koda od 64 ne slu?e kao kod za aminokiselinu. Oni ozna?avaju kraj sinteze proteina i nazivaju se terminiraju?im ili besmislenim kodonima (stop kodon). Njihovo postojanje je bilo
Univerzalnost koda
Zna?enje kodona je razja?njeno tokom eksperimenata na bakteriji E. coli. Ali ?ta ako geni drugih organizama, uklju?uju?i ljude, koriste druga?iju ?ifru? U ovom slu?aju, mutacije u ljudskim proteinima
Nasljednostb u svijetu bakterija
Predstavnici klasi?ne genetike jedva su se usudili sanjati o mogu?nostima koje se otvaraju pred modernim znanstvenicima koji sprovode eksperimente na bakterijama i virusima bakterija. U ovom poglavlju mi
Mutantne bakterije
Razli?ite vrste bakterija mogu se razlikovati po fenotipskim karakteristikama, kao ?to su oblik, boja i drugi karakteristi?ni detalji njihovih kolonija. Ali u toku istra?ivanja napravljen je veliki napredak u genetici bakterija.
Seks u E. coli
Godine 1946. Joshua Lederberg i Edward Tatham po?eli su provoditi genetske eksperimente na bakterijama. Nekoliko godina ranije, Tatem je radio sa George Beall-om, i to na osnovu op.
Plazmidi
Faktor F je primjer takozvanog plazmida, odnosno ekstrahromozomskog samorepliciraju?eg genetskog elementa kru?ne strukture. Plazmidi su svojevrsni putnici u ?eliji, koji
Faktori rezistencije i rezistencija na antibiotike
1955. godine, Japanka se vratila iz Hong Konga sa oblikom dizenterije izazvanom bakterijom iz roda Shigella. Shigella infekcija se lako lije?i antibioticima, ali ove bakterije
Lizogenija
Biolozi koji su eksperimentirali s fagima prije Drugog svjetskog rata ?esto su tvrdili da odre?eni sojevi bakterija nose viruse koji se ponekad nepredvidivo manifestiraju u kulturama koje rastu.
Geni koje prenosi virus
Poku?avaju?i utvrditi konjugate salmonele poput E. coli, Norton Zinder je otkrio da fagi mogu prenijeti gene iz jedne bakterijske ?elije u drugu. Ovaj fenomen se naziva t
Transdukcija i ljudski genom
Joshua Lederberg je 1955. godine predlo?io da se transduciraju?i virusi mogu koristiti za uvo?enje gena u ljudske ?elije. U to vrijeme takva ideja je izgledala kao ?ista fantazija, ali sada je sve vi?e
Regulacija gena i razvoj organizma
Dok ?itamo knjigu, kao i kroz istoriju genetike, na?e razumevanje gena se stalno menjalo. Ako smo u po?etku gen smatrali neodre?enim faktorom to nekako prije
Regulacija gena u bakterijama
Kao iu prethodnim poglavljima, po?e?emo od jednostavnih biolo?kih sistema, odnosno od bakterija, u ?ijem prou?avanju se ovo pitanje prvi put postavilo. Istra?ivanja su vr?ena uglavnom 1950-ih i 1960-ih godina.
Regulacija eukariotskih gena
Pitanje regulacije gena u eukariotskim stanicama zahtijeva druga?iju formulaciju, budu?i da se na?in ?ivota tipi?nih eukariota bitno razlikuje od na?ina ?ivota prokariota. Prokarioti su jednostavne bakterije
Embrionalni razvoj op?enito
Embrion se razvija iz jedne ?elije - zigota - i pretvara se u kompleks mnogih specijalizovanih ?elija. Zigota je totipotentna, odnosno nakon ponovljene diobe mo?e datirati
Vremenska regulacija i razvoj krila pili?a
Odli?an primjer mehanizma za mjerenje vremena je razvoj krila pili?a (slika 11.2). Krilo raste iz dodatka udova, koje se sastoji od ?elija mezoderma prekrivenih slojem ektoderma, uklju?uju?i apikalni region.
Formiranje oka muhe
Jedan od najzanimljivijih serijala doga?aja koji uklju?uje vi?e gena doga?a se tokom formiranja oka Drosophile. Slo?eno oko insekta sastoji se od otprilike 800 elemenata. Odvojeni element
Intervencija u strukturi DNK: povratak Epimeteja?
U drevnim gr?kim mitovima govorilo se o titanima - rasi divova koju su rodili bogovi prije rase ljudi. Bogovi su uputili titana Epimeteja da stvori ?ivotinje i biljke, dijele?i me?u njima razne
Rekombinantna DNK i restrikcijski enzimi
Do 1972. Annie Chang, Paul Berg i Seymour Cohen su ustanovili da restrikcijski enzimi mogu presje?i bilo koja dva molekula DNK i od njih napraviti jednu rekombinantnu DNK.
Ispitivanje pojedina?nih kloniranih fragmenata
?esto je pa?nja eksperimentatora usmjerena na jedan gen donora koji nau?nici ?ele prou?iti ili koristiti. Uz malo sre?e, takav gen mo?e ve? biti sadr?an u genomu
Genska terapija
Me?u razli?itim na?inima kori?tenja transgenih tehnologija posebno mjesto zauzima genska terapija. Ako je mogu?e modificirati biljne i ?ivotinjske organizme, ?ta onda sprje?ava primjenu istih
Genomika - prou?avanje ?itavog genoma
Nedavni napredak u sekvenciranju i razvoju tehni?kih sredstava za obradu velikog broja klonova u biblioteci gena omogu?ili su nau?nicima da prou?e cijeli genom organizma odjednom. Se
Geneti?ar kao dr Franken?tajn
U o?ima moderne javnosti, geneti?ari se ?esto povezuju sa slikom junaka romana Mary Shelley "Frankenstein", ludo strastvenog prema svom poslu i stvaranju stra?nog ?udovi?ta. Krivim geneti?are
Kontrola istra?ivanja rekombinantne DNK
Debata o ulozi genetike zapo?ela je mnogo prije modernog procvata genetskog in?enjeringa. Jo? 1970-ih godina, ne samo nau?na zajednica, ve? i ?ira javnost po?ela je da raspravlja o pitanjima vezanim za
genetski modifikovanih organizama
Pitanja uticaja javnosti na genetiku i regulisanje nau?nih istra?ivanja u ovoj oblasti jo? uvek u mnogome nisu re?ena. Kako se mikrobiolo?ke tehnologije i metode pobolj?avaju,
Tehnologija u kontekstu
Jedna od strana problema koji se pojavio je nau?no prosvjetljenje. Kako se ?ale agenti za nekretnine, tri klju?na elementa za prodaju ku?e su lokacija, lokacija i lokacija. Upravo isto
Argumenti protiv genetski modifikovane hrane
Tokom kontroverze oko genetski modifikovane hrane, izneti su brojni argumenti protiv njihove upotrebe. Ovdje navodimo glavne argumente protivnika koji su u osnovi njihovog razmi?ljanja.
Eti?ki aspekti kloniranja
Kloniranje ?ivotinja, iako nije direktno povezano s transgenim tehnologijama, tako?er postavlja sli?na eti?ka pitanja. Prije svega, to se ti?e sisara. O tome se odavno zna
Odgovornost nau?nika
Moderne genetske tehnologije sposobne su nanijeti zna?ajnu ?tetu ?ovje?anstvu, pa stoga dru?tvo mora stalno biti na oprezu. Na? cilj nije ?tititi ili osu?ivati genetske tehnologije.
Frekvencija mutacije
Mutacije se uvijek de?avaju prirodno, nasumi?no i bez vidljivog razloga. Ne mo?emo unaprijed predvidjeti koja ?e se mutacija dogoditi i gdje se, stoga, pri njihovom prou?avanju koriste statisti?ke metode.
Mutacije kod ljudi
Stope mutacija kod ljudi mogu se odrediti kori?tenjem pedigrea koji pokazuju dominantne osobine. Defekt koji se iznenada pojavio kod jednog predstavnika generacije i pre?ao na potomstvo
Radijacija
Spontane mutacije su prili?no rijetke. U?estalost mutacija pove?avaju mutageni. Najmo?niji mutageni uklju?uju odre?ene vrste zra?enja. Godine 1927. Hermann M?ller, eksperimentiraju?i s Drosophila, i
?ta su mutacije?
Mutacija je promjena u DNK. Neke promene se de?avaju spontano, tokom vremena. Na primjer, molekuli DNK gube purinske baze guanin i adenin (depurinacija) relativno velikom brzinom.
Sistem za popravku DNK
Kako se ?ivot razvijao na na?oj planeti, ?elije su se stalno susrele sa raznim mutagenima, kako u obliku zra?enja tako i u obliku hemikalija. Stopa mutacije mora biti unutar
Genetske posljedice zra?enja
Jonizuju?e zra?enje uzrokuje mutacije bilo koje vrste - od zamjenskih ta?aka do kromosomskih aberacija i lomova. Postavljanjem izvora niske radijacije u ?umu, istra?iva?i su dokazali da je stalna radijacija
Hromozomske aberacije
Hromozomi sadr?e gene raspore?ene u odre?enom nizu. Fenotip organizma ne zavisi samo od odre?enih gena, ve? i od toga kako se nalaze u odnosu na druge gene. Na
ljudski hromozomi
Pod elektronskim mikroskopom, ljudski hromozomi izgledaju kao komadi?i debelog u?eta upletenog u brojne petlje. Svaki hromozom je duga?ak, kontinuirani lanac DNK, umotan
Aneuploidija
Bogat izvor materijala za prou?avanje hromozomskih aberacija su poba?aji u prvim nedeljama razvoja, jer imaju 50-100 puta vi?e hromozomskih abnormalnosti nego novoro?en?e
umno?avanje i brisanje
Duplikacije i delecije velikih dijelova hromozoma su gotovo uvijek smrtonosne, kao i ve?ina mutacija. Ako fetus pre?ivi, onda ga karakteriziraju ozbiljni poreme?aji u razvoju. Najpoznatiji premijer
Inverzije
Mnogi od nas su ?uli za parove koji ne uspijevaju imati djecu zbog poba?aja i poba?aja. Ovo se doga?a ako je jedan od partnera heterozigotan za inverziju ili translokaciju.
Translokacije
Translokacije su ?est uzrok nasljednih poreme?aja koji se mogu vidjeti u kariotipu. Obi?no ih nose heterozigoti koji imaju jedan normalan kromosom i jedan kromosom s translokacijom.
Dokazi za evoluciju
Dokazi da su razli?iti organizmi zapravo evoluirali od zajedni?kog pretka kroz postepenu promjenu dolaze iz razli?itih izvora. Mo?da jedan od najja?ih dokaza
Evolucija kao proces
U ?irem smislu, evolucija obuhvata tri procesa: makroevoluciju, specijalizaciju i mikroevoluciju. Makroevolucija podrazumijeva ukupnost svih procesa zbog kojih su u pro?losti bi?a
populaciona genetika
Podjela alela gena na divlje i mutantne, kao ?to smo radili prilikom upoznavanja sa osnovama genetike, nije sasvim ispravna, a takva podjela mo?e dovesti do pogre?ne predstave o evoluciji. istra?ivao
ljudska evolucija
Najkontroverzniji zaklju?ak iz Darwinove teorije evolucije bila je pretpostavka da je ?ovjek evoluirao od majmuna. Predstavnici hri??anske religije su ovu ideju prihvatili sa neodobravanjem, jer
Migracija i raznolikost Homo sapiensa
Zahvaljuju?i DNK sekvenciranju ljudi ?irom svijeta, bilo je mogu?e izgraditi filogenetsko stablo ?ovje?anstva. Korijenje ovog drveta, kako svjedo?e fosilizirani ostaci, ide u Afriku. Most bio
Boja ko?e
Prosje?na boja ko?e populacije gotovo je direktno povezana s geografskom du?inom: najtamniji se nalazi u blizini ekvatora, a najsvjetliji je bli?e polovima. Tamnija ko?a pru?a bolju UV za?titu.
Eugenika
Kao ?to je ve? pomenuto u pogl. 1, ideja o pobolj?anju ljudske rase nastala je davno, barem u drevnom gr?kom dru?tvu klasi?nog perioda. Ali je privukao posebnu pa?nju posljednjih godina.
Dictionaryb
Autoradiografija je metoda dobijanja slike radioaktivnih materijala izlaganjem fotografskom rastvoru; gdje se pojavljuje otopina formira se tamna mrlja.
Karboksilna grupa - hemijsko jedinjenje COOH, koje se naziva i kiselinska grupa, jer atom vodonika te?i da se odvoji u obliku H+ jona
Karboksi kraj - kraj polipeptidnog lanca sa slobodnom karboksilnom grupom. Kariotip je dijagram hromozomskog seta organizma koji je rezultat
Citozin je jedna od pirimidinskih baza DNK ili RNK
Frekvencija alela - u populacionoj genetici, odnos nekoliko alela jednog gena (ili tipa hromozoma). Stopa mutacije je mjera vjerovatno?e da a






